3 Cascade Pathways to Generate C3 Convertase
1. Classical Pathway: Innate & Adaptive Immunity
The classical pathway begins primarily when two or more antigen-antibody complexes (immune complexes) bind to the complement C1 complex. Since it involves antibody binding, this pathway can be attributed to adaptive immunity. Among the five classes of antibodies (Ig: Immunoglobulin), only those of the IgM and IgG subclasses can activate the complement by binding to it. However, this pathway can also be initiated by directly binding to certain pathogens, contributing to innate immunity. Components such as C-reactive protein (CRP), amyloid-beta peptides (which can form amyloid plaques and contribute to Alzheimer's disease), damaged tissue fragments, aged cells, and parts of cells undergoing or improperly undergoing apoptosis (like DNA and RNA) are recognized and bound by the complement, ensuring its activation. These components are referred to as pattern recognition elements. Particularly, recognizing and timely removing host cells or tissues that need to be eliminated is very important as it is related to diseases such as autoimmunity. However, generally, the classical pathway refers to the initiation of activation by binding to antibodies.
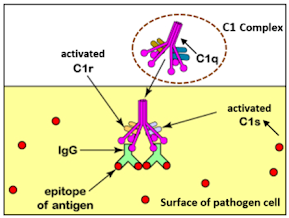
C1 complex activation
When observed under a microscope, the first component of the complement system, C1, appears somewhat like an upside-down bunch of tulips or a frog's webbed foot. C1 is a complex consisting of C1q, C1r, and C1s, where C1r and C1s function as serine proteases (protein-degrading enzymes). C1q (depicted in pink in the diagram below) is a type of collectin (collagen + lectin) protein, composed of a rod-like part with collagen-like domains and a round head part with lectin-like domains.
When the head portion of C1q binds to two or more antibodies (specifically the Fc region) or directly to pathogens, it undergoes a structural change that activates the C1r enzyme. The activated C1r then cleaves and activates C1s. In this process, C1s transitions from its zymogen (inactive enzyme precursor) state to an activated enzyme.
C4, C2 activation
Now, C1s cleaves the complement molecules C4 and C2 into C4a, C4b, and C2a, C2b, respectively. Remember that one molecule of C1s can cleave multiple C4 and C2 molecules. The complex formed by C2a and C4b will become the C3 convertase. Let's examine this step by step.
First, C1s cleaves C4, and the small fragment C4a initiates a localized inflammatory response, although its effect is minimal. Once C4 is cleaved, the thioester bond within C4b is exposed, allowing it to react and covalently attach to the surface of pathogens. Then, C2 binds to the C4b that is attached to the pathogen's surface. In this bound state, it becomes easier for C1s to cleave and activate C2.
C2 is then cleaved into C2a and C2b (as mentioned previously, only in case of C2, the bigger fragment is named “a”). The small fragment C2b is released, while the C2a remains attached to the pathogen's cell surface together with C4b, forming the complex known as the C3 convertase. The C3 convertase is generated. In this C3 convertase complex, the C2a fragment serves as the activated serine protease that primarily cleaves the C3 molecule.
It may have been clearer to refer to it as the C3 cleaving enzyme rather than the C3 convertase (I always have complaints about naming conventions, haha). The C3 convertase, specifically the C4b2a complex, now actively cleaves nearby C3 molecules, with a single C convertase capable of cleaving and activating up to 1,000 C3 molecules.
2. Lectin pathway : innate immunity
The lectin pathway is very similar to the classical pathway but has some differences. While the classical pathway primarily recognizes pathogens under the leadership of antibodies, the lectin pathway uses lectins—soluble proteins capable of recognizing specific carbohydrates—as receptors to identify antigens that arise from the external environment. This pathway operates independently of antibodies and is thus classified under the innate immune system.
2-1 Mannose-binding lectin (MBL) pathway
The activation occurs through MBL, which binds to mannose, instead of the C1 complex. This pathway is quite similar to the classical complement activation process. The difference lies in the fact that MBL performs the function of the C1 complex, while the roles of C1r and C1s are taken on by MASP-1 and MASP-2. MASP, which stands for MBL-associated serine protease, activates and cleaves C4 and C2. Like the C1 complex, MBL also contains collagen-like domains and lectin-binding domains (carbohydrate recognition sites), classifying it within the collectin family.
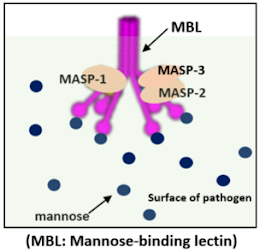
How MBL Recognizes Pathogens
Mannose-Binding Lectin (MBL) can recognize and bind to pathogens even in the absence of antibodies. Our cells are surrounded by a cell membrane made of two layers of phospholipids. Additionally, glycoproteins, which can either protrude outward or pass through the membrane, exist on the surface of the cell membrane. Glycoproteins consist of proteins covalently bonded to oligosaccharides (short polysaccharides), with most of their terminal residues being sialic acid or galactose.
However, the glycoproteins of yeasts are predominantly composed of the monosaccharide mannose, and mannose can also be found in the cell walls of bacteria or some viruses. For example, mannose residues are present on the surfaces of microorganisms like Salmonella and Listeria, as well as on the cell surfaces of fungi like Candida and viruses such as HIV-1. This means that the terminal carbohydrate components differ from those of the host.
Pattern recognition receptors (PRRs) such as collectins have structures that allow them to identify patterns that are distinct from the carbohydrate terminal patterns of "self". When they repeatedly encounter differently shaped patterns, they recognize them as foreign. Because PRRs can distinguish between self and non-self carbohydrates and bind to various microorganisms to perform necessary effector functions, they are considered a primary defense mechanism of the innate immune system. MBL is produced in the liver and exists at low concentrations in the bloodstream; however, its production dramatically increases as part of the acute phase response during infection
 |
In the classical pathway, when C1q binds to antibodies or pathogens, C1r is automatically activated, which in turn activates C1s. Similarly, when MBL (Mannose-Binding Lectin) binds to a pathogen, MASP-1 (MBL-associated serine protease-1) is auto-activated, leading to a structural change, followed shortly by the cleavage and activation of MASP-2. Both MASP-1 and MASP-2 exist as inactive precursors (zymogens) and are activated consequently. Following this, they undergo the same cleavage process as in the classical pathway to generate the C3 convertase. MASP-1 and MASP-2 both cleave C2, but only MASP-2 cleaves C4. The catalytic activity for generating C3 convertase is reported to be 1000 times higher for MASP-2 compared to C1 in the classical pathway.[1] This allows the innate immune system to buy time until antibodies can be produced.
MASP-3 has a competitive relationship with MASP-2, potentially decreasing the synthesis of C3 convertase, thus acting as an inhibitor of complement action. However, there is a relatively recentl study(2016) reporting that MASP-3 can exclusively activate complement factor D, which is crucial for cleaving and activating complement factor B in the alternative pathway, indicating that the lectin pathway and the alternative pathway are fundamentally interconnected.[2]
2-2 Ficolin Pathway
Ficolin is another type of pattern recognition receptor utilized in the lectin pathway. Ficolin is very similar to MBL in that both MBL and ficolin possess collagen-like domains; however, while MBL has a lectin-binding domain, ficolin instead has a fibrinogen-like domain. Ficolin is actually referred to as "a new form of lectin that uses fibrinogen-like domains as functional domains" [3].
Due to its fibrinogen-like domain, ficolin exhibits specificity for acetylated oligosaccharides found on bacteria and does not bind to carbohydrates with mannose residues. The human body has three types of ficolin: L-ficolin (ficolin-2), M-ficolin (ficolin-1), and H-ficolin (ficolin-3). L-ficolin and H-ficolin are synthesized in the liver and circulate in the blood, while M-ficolin is secreted from the lungs and blood cells.
The ficolin pathway, similar to the MBL pathway, progresses to generate the C3 convertase through the activation of MASP-1 and MASP-2 in the same manner.
3. Alternative pathway: innate immunity
To initiate the cascade activation steps of the classical or lectin pathways, they must first bind to antibodies or to external substances such as carbohydrates on the surface of pathogens. However, the alternative pathway generates C3 convertase without this direct binding process. The alternative pathway has two starting points: 1. Using C3b: The first begins with C3b that has already been generated through the previously known 2 pathways. 2. Self-Hydrolysis of C3: The second begins when the unstable C3 undergoes spontaneous hydrolysis to form C3(H2O). This spontaneous hydrolysis is referred to as the "tick over" process, which is a phenomenon unique to the alternative pathway.
There are complement components that appear exclusively in the alternative pathway: Complement factor B (CFB), Complement factor D (CFD), and Properdin (Complement factor P, CFP). Let's explore their roles in the process of the alternative pathway.
Generation of C3 Convertase from Pre-existing C3b
First, let's examine how the alternative pathway generates C3 convertase using existing C3b fragments. After being generated through the classical or lectin pathways, the C3b fragment, which is covalently bonded to the surface of pathogens like microorganisms, binds to complement factor B. This binding induces a structural change in factor B, allowing the serum protease, another complement factor D, to easily cleave factor B into Ba and Bb fragments. Factor D is known to circulate in its active form and is unique in that its function corresponds to elements not found in the classical or lectin pathways.
Factor B is closely related to C2 in the classical pathway, showing structural and functional similarities, and both exist as inactive zymogens. After cleavage, Ba dissociates, leaving Bb bound to the C3b fragment on the cell surface, thus forming the complex (C3bBb), which acts as C3 convertase.
"Tick over" process
Now, let's examine the alternative pathway through a spontaneous process. The thiol ester bond within C3 reacts with surrounding water, leading to hydrolysis, or it interacts with small amounts of plasma proteolytic enzymes, resulting in the formation of a structurally altered intermediate known as C3(H2O) or iC3. This phenomenon, where approximately 1% of the total C3 molecules undergoes a very slow but steady spontaneous hydrolysis, is referred to as the "tick over" process. In this situation, in the presence of magnesium (Mg²+), complement factor B binds to the spontaneously generated C3(H2O).
Similarly, factor B bound to C3(H2O) can also be cleaved by complement factor D, yielding Ba and Bb fragments. The Bb fragment, together with C3(H2O), forms a complex (C3(H2O)Bb), which acts as C3 convertase. These enzymes remain in serum but are not attached to the surface of pathogen cells, thus this step is referred to as fluid-phase convertase. This convertase has a short lifespan in serum but can cleave multiple C3 molecules into C3a and C3b very quickly.
The produced C3b can then bind again to complement factor B, replacing the C3(H2O)Bb fluid-phase convertase with C3bBb C3 convertase in a manner similar to the first method of starting the alternative pathway. However, C3bBb C3 convertase is still unstable; thus, Properdin (or Complement factor P) binds to it to enhance its stability. Properdin is a component released in the granules of activated neutrophils or macrophages for the immune response and can bind to the surface of pathogens, thereby stabilizing the newly formed C3 convertases on the cell surface and preventing their degradation.
With the enhanced stability provided by Properdin, the C3bBb C3 convertase can quickly generate C3b molecules. This process can then repeat, forming a positive feedback loop that produces a tremendous number of C3b in a short period.
In summary, the alternative pathway includes two forms of the C3 convertase: the C3(H2O)Bb C3 convertase (fluid phase), which is generated through spontaneous hydrolysis and exists in serum, and the C3bBb C3 convertase, formed when C3b fragment generated by this fluid phase convertase or through 2 other pathways combines with the Bb fragment. A key feature of this pathway is that it can amplify the production of C3b through positive feedback. Now we will see that C3b is directly involved in the generation of the C5 convertase.

C5 Convertase
We have examined the three pathways that produce C3 convertase in detail. Now, all these pathways converge to bind the C3b fragment generated by their respective C3 convertases to form the C5 convertase, which subsequently cleaves C5 into C5a and C5b.

Having explored how these fragments are cleaved, let’s refer to another diagram that illustrates the entire cascade process of complement activation, starting from the three initial pathways to the final step of forming the Membrane Attack Complex (MAC). Now, we will examine the detailed functions of the complement system in the next article.

[References]
[1] MASP1 and MASP2
https://doi.org/10.1007/978-1-4614-6438-9_101714-1
[2] MASP-3 is the exclusive pro-factor D activator in resting blood: the lectin and the alternative complement pathways are fundamentally linked
https://doi.org/10.1038/srep31877
[3] The role of ficolins in the lectin pathway of innate immunity
https://doi.org/10.1016/j.biocel.2011.02.003
[4] Roitt's Essential Immunology, Thirteenth Edition. Peter J. Delves, Seamus J. Martin, Dennis R. Burton, and Ivan M. Roitt. Published 2017 by John Wiley & Sons, Ltd
[Basic readings]
Kuby Immunology. 8th New York: Macmillan Learning, 2019. Text. MLA Style. Punt, Jenni, Stranford, Sharon A, Jones, Patricia P, Owen, Judith A.
Janeway's immunobiology. Kenneth Murphy, Janeway Jr., Paul Travers, Walport Sir. 9th Edition, New York, Garland Science/Taylor & Francis Group, LLC, [2016]
Fundamental Immunology 5th edition (August 2003): William E. Paul (Editor). Philadelphia: Lippincott Williams & Wilkins, c2003.


